Eón Proterozoico
2.500 MILLONES DE AÑOS
EÓN PROTEROZOICO
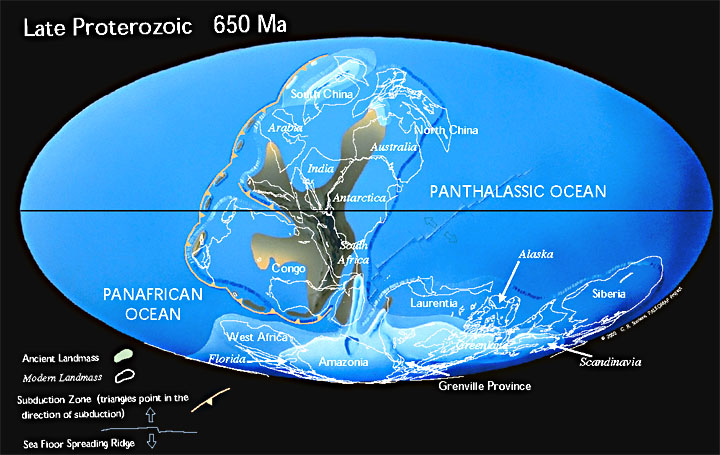
El Proterozoico (de próteros = anterior, temprano y zôon = ser vivo), una división de la escala temporal geológica antes también conocida como Algónquico o Eozoico, es un eón geológico perteneciente al Precámbrico que abarca desde hace 2500 millones de años hasta hace 542 millones de años, durando 1958 ± 1,0 millones de años. Se caracteriza por la presencia de grandes cratones que darán lugar a las plataformas continentales. Las cordilleras generadas en este eón sufrieron los mismos procesos que los fanerozoicos. La intensidad del metamorfismo disminuyó en este momento geológico. La Tierra sufre sus primeras glaciaciones y se registra una gran cantidad de estromatolitos. Las primeras glaciaciones conocidas se produjeron durante el Proterozoico. La primera, la Glaciación Huroniana, se produjo poco después del comienzo del eón en el Período Riásico y culminaron en el Período Criogénico con la hipótesis de la Tierra bola de nieve. Sin duda, supusieron un importante cambio en la biota terrestre. El período Ediacárico de finales del Proterozoico se caracteriza por la evolución de abundantes organismos pluricelulares de cuerpo blando.
ERA PALEOPROTEROZOICA

Orógeno Wopmay
El Paleoproterozoico es una división de la escala temporal geológica, es la primera era geológica de las tres que componen el eón proteozoico y que comienza hace 2.500 millones de años y termina hace 1.600 millones de años durando 900 millones de años. Es en esta era cuando los continentes se estabilizaron por primera vez. También se produjo la Gran Oxidación como consecuencia del proceso químico de fotosíntesis realizada por las cianobacterias. Durante esta época aparecen los primeros cinturones montañosos que sobreviven en la actualidad, como el Orógeno Wopmay de Canadá (de hace 2.100-1.800 millones de años).
Durante esta era se generan las primeras montañas a través de procesos parecidos a los fanerozoicos, aproximadamente hace 2.100-1.800 millones de años, en el Orógeno de Wopmay (Canadá), al oeste de la Bahía de Hudson. La secuencia de depósitos formados es la siguiente:
Areniscas cuarcíticas
Dolomías con estromatolitos y depósitos mareales y de laguna.
Lutitas de aguas cada vez más profundas.
Depósitos de tipo flysh (turbiditas) de aguas profundas.
Turbiditas carbonatadas en transición a facies con estromatolitos y grietas de desecación (medios profundos a someros).
Depósitos de molasas fluviales (materiales postorogénicos generalmente discordantes).
PERIODO SIDÉRICO

El período Sidérico (2.500 – 2.300 Ma) es la primera de las subdivisiones de la era paleoprotozoica; está caracterizado por la fragmentación del Kenorland y el nacimiento del continente Ártica, por el inicio del depósito de las formaciones de hierro bandeado, resultado de la contaminación atmosférica por parte del oxígeno, y por el comienzo de una gigantesca glaciación (ocurrida hace unos 2.400 Ma) que cubriría el globo terráqueo: la glaciación Huroniana.
La cantidad de oxígeno que se fue liberando por acción de los organismos fotosintéticos (estromatolitos) fue aumentando poco a poco, llegando a acumularse en tales cantidades que dio inicio, hace unos 2.450 Ma, a la oxidación del hierro disuelto en las aguas de los océanos, generándose grandes cantidades de Fe3O4(magnetita), que se fueron depositando en los fondos marinos
Este proceso fue eliminando el hierro disuelto de los océanos, que poco a poco fueron perdiendo su coloración verdosa y adquiriendo un aspecto más azulado; una vez saturados los océanos de oxígeno, este empezó a extenderse por la atmósfera. A este evento se lo conoce como “la Gran Oxidación”
2.400 MILLONES DE AÑOS
LA GRAN OXIDACIÓN

Hierro bandeado
La Gran Oxidación fue un cambio medioambiental muy importante que ocurrió, probablemente, durante el periodo Sidérico, (hace unos 2.400 Ma), y que tuvo como resultado el depósito de una gran cantidad de hierro oxidado, que hasta entonces había permanecido disuelto en las aguas oceánicas. Dada la importancia del oxígeno en este cambio medioambiental, a este evento también se lo suele denominar crisis del oxígeno , revolución del oxígeno o catástrofe del oxígeno .
Los primeros organismos fotosintéticos que existieron realizaban la fotosíntesis anoxigénica, en la cual no se desprende oxígeno (tal y como hacen en la actualidad las bacterias verdes del azufre o las bacterias púrpura). Pero cuando surgieron los primeros organismos capaces de realizar la fotosíntesis oxigénica (como las cianobacterias), se empezó a producir oxígeno molecular (O2) en grandes cantidades, que fue lentamente invadiendo la atmósfera y los océanos. La emisión de tal exceso de oxígeno al medio ambiente provocó una crisis ecológica de la época, pues el oxígeno era tóxico para los microorganismos anaerobios dominantes por aquel entonces.
Mientras esto ocurría, el oxígeno que se iba produciendo se consumía en diversas reacciones químicas que tenían lugar en los océanos, pues reaccionaba principalmente con el hierro y generaba minerales tales como la magnetita (Fe3O4) o el hematites (Fe2O3), que precipitaban y se depositaban en los fondos oceánicos y de aguas continentales. El agua, que hasta aquel entonces había sido de color verde debido al hierro disuelto en ella, empezó a perder este hierro, lo que le llevaría a adoptar su coloración azulada actual. La prueba de este fenómeno se encuentra en las antiguas rocas que contienen enormes formaciones de hierro bandeado o BIF , procedentes de la oxidación del hierro presente en el océano y que en la actualidad constituyen los principales yacimientos de hierro explotados comercialmente.

Magnetita
Otra consecuencia importante fueron los cambios climáticos subsiguientes. La generación demetano (CH4) atmosférico se debía en buena parte a los organismos anaerobios, los cuales sufrieron descensos poblacionales debido al aumento del oxígeno de la atmósfera. Por otro lado, el metano, frente al oxígeno molecular y la radiación ultravioleta, se oxida rápidamente, generando dióxido de carbono (CO2). Este cambio de CH4 a CO2 en la atmósfera reduciría de forma considerable la temperatura global, ya que el potencial de efecto invernadero del metano es varias veces mayor que el del dióxido de carbono. Este descenso drástico de la temperatura desencadenaría la glaciación Huroniana, ocurrida hace 2.400 – 2.100 Ma aproximadamente.

Hematites
Sin embargo, esta drástica transformación también ofreció una nueva oportunidad para la diversificación biológica, así como enormes cambios en la naturaleza de las interacciones químicas entre las rocas, la arena, las arcillas y otros sustratos geológicos, y la atmósfera, los océanos y otras aguas superficiales. A pesar del reciclado natural de la materia orgánica, la vida se había mantenido energéticamente limitada hasta la amplia disponibilidad de oxígeno. Este avance en la evolución del metabolismo aumentó en gran medida el suministro de energía para los organismos vivos, produciendo un impacto ambiental global.
MORALEJA
Sin duda alguna las protagonistas de nuestra historia son las cianobacterias,sin ellas no se hubiera liberado oxigeno,esta liberación de oxigeno en la atmósfera dio lugar a otras formas de vida que fueron evolucionando.
GLACIACIÓN HURONIANA

La glaciación Huroniana fue una hipotética glaciación ocurrida hace unos 2.400 – 2.100 Ma, durante los períodos Sidérico y Riásico. Según las interpretaciones de numerosos geólogos y glaciólogos, esta glaciación habría sido una de las más intensas de todo el registro geológico, llegando a tener unos efectos muy similares a los de la glaciación Sturtiense-Varangiense (que ocurrió a finales del eón Proterozoico y desarrolló un estado de Tierra “bola de nieve”).
A comienzos de la glaciación Huroniana existían varias masas continentales principales formadas a partir de la lenta fragmentación del gigantesco supercontinente Kenorland (Ur,Ártica,Báltica y los restos de Kenorland).
La apertura de nuevos océanos empujaba a todos estos continentes, provocando la colisión de unos cratones con otros y generando una intensa etapa orogenética. Así mismo, al fragmentarse Kenorland en varias masas continentales más pequeñas, la superficie continental invadida por el mar se incrementó, generándose multitud de mares someros y permitiendo un rápido desarrollo y expansión de las plataformas continentales, lo que a su vez favoreció el desarrollo de extensas comunidades de tapices microbianos productores de oxígeno.
Aunque a día de hoy todavía no hay consenso sobre las causas que llevaron a la glaciación Huroniana, se cree que la causa principal de que se produjera esta glaciación fue la intensa desestabilización del clima provocada por el metabolismo de las primeras cianobacterias (que ya habían desencadenado, en el Sidérico, la Gran Oxidación), al romper el equilibrio existente entre los gases de efecto invernadero por adición de oxígeno y eliminación del CO2 durante la realización de la fotosíntesis oxigénica, y por la oxidación de metano a dióxido de carbono en contacto con el oxígeno atmosférico.
Del mismo modo que no se sabe con certeza qué provocó la glaciación Huroniana, se desconocen cuáles fueron las causas que llevaron al fin de esta glaciación, siendo muy probable que su fin se debiera al intenso vulcanismo que estaba teniendo lugar durante la fragmentación de Kenorland y a la desaparición de la gran parte de los estromatolitos provocada por la propia glaciación, lo que acabaría con la retirada masiva del CO2 de la atmósfera, y permitiría que tanto éste como el metano se volviesen a acumular en ella.
2.300 MILLONES DE AÑOS
PERIODO RIÁSICO

Complejo ígneo de Bushveld
El período Riásico (2.300 – 2.050 Ma), también llamado Riácico o Riaciense, es la segunda de las subdivisiones de la era Paleoprotozoica; está caracterizado por la gigantesca glaciación Huroniana (que empezó hace 2.400 Ma), durante la cual el globo terráqueo podría haber quedado cubierto de hielo por –o casi por– completo (hasta que la glaciación terminó de forma abrupta hace unos 2.100 Ma), y por el inicio localizado de varias etapas orogénicas: la orogenia Ebúrnica(en el cratón de África Occidental, hace unos 2.200 Ma), la orogenia Wopmay (hace 2.100 Ma) y la orogenia Buganda-Toro-Kibaliana (hace 2.075 Ma), que dieron lugar a la formación de un buen número de complejos ígneos (como el complejo ígneo de Bushveld, en Sudáfrica).
2.200 MILLONES DE AÑOS
TEORÍA ENDOSIMBIÓTICA
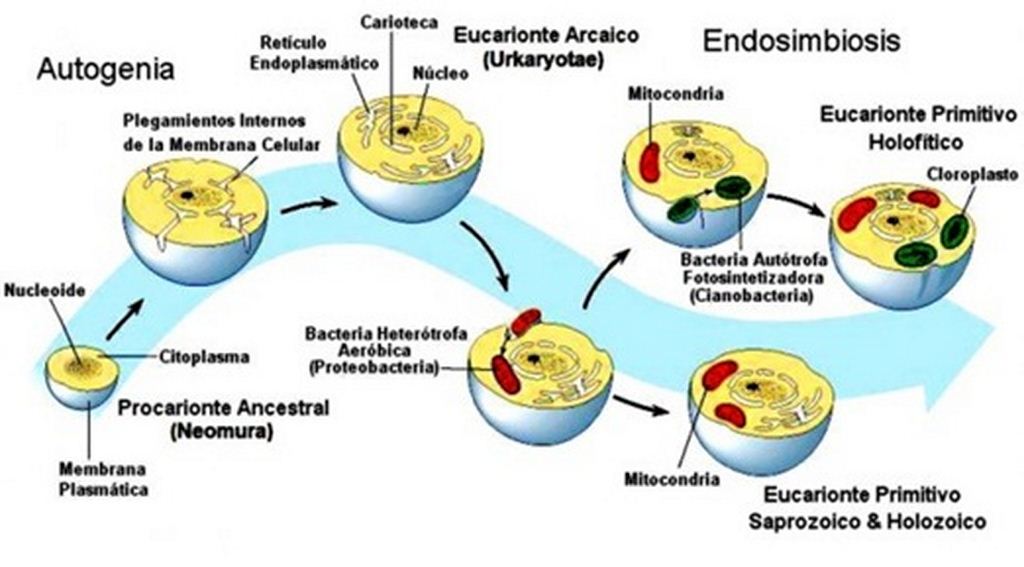
Se llama endosimbiosis a la asociación de un organismo que vive dentro de otro organismo. Lynn Margulis propuso la teoría de la endosimbiosis seriada, que explica la aparición de la célula eucariótica por asimilación simbiótica de varias bacterias con habilidades diferentes.
-
- 1.Fusión de dos procariontes: uno irregular o redondeado, que utilizaba el azufre y el calor como fuente de energía, y otro nadador, similar a espiroquetas. Surgiendo un tipo de organismo anaeróbico, en el que el ADN de ambos procariontes habría quedado encerrado dentro de una membrana.
-
- 2.El nuevo organismo, con núcleo y anaeróbico, incorporó un procarionte que podía vivir en ambientes con oxígeno que crearon las mitocondrias. Así la célula con núcleo adquirió la capacidad de vivir en medios aerobios. Esto les proporcionó a ambos organismos una ventaja respecto de los procariontes anaeróbicos, y la célula hospedadora, en lugar de digerir al procarionte incorporado, estableció con él una relación simbiótica en la que uno proporciona nutrientes, y el otro energía suficiente para ambos. Así nacen las células eucariotas con mitocondrias, de nutrición heterótrofa que originaría a las primeras células animales.
-
- 3.Las células con núcleo, heterótrofas y aeróbicas (con mitocondrias), incorporaron procariontes de vida libre fotosintéticos que sintetizaban materia orgánica gracias a la energía del Sol. Así nacen las células con cloroplastos (los que producen la fotosíntesis), de nutrición autótrofa que tenían autonomía al producir su propio alimento. Es decir aparecen las primeras células eucariontes vegetales.
Esta teoría no se pudo dar en su totalidad hasta hace 1.500 millones de años ,pero hace 2.200 Ma se supone que aparecieron las mitocondrias pero no la célula eucariota en si.
Mitocondria
* El tamaño de las mitocondrias es similar al tamaño de algunas bacterias.
* Las mitocondria y los cloroplastos contienen ADN bicatenario circular cerrado covalentemente – al igual que los procariotas- mientras que el núcleo eucariota posee varios cromosomas bicatenarios lineales.
* Están rodeados por una doble membrana, lo que concuerda con la idea de la fagocitosis: la membrana interna sería la membrana plasmática originaria de la bacteria, mientras que la membrana externa correspondería a aquella porción que la habría englobado en una vesícula.
* Las mitocondrias y los cloroplastos se dividen por fisión binaria al igual que los procariotas (los eucariotas lo hacen por mitosis). En algunas algas, tales como Euglena, los plastos pueden ser destruidos por ciertos productos químicos o la ausencia prolongada de luz sin que el resto de la célula se vea afectada. En estos casos, los plastos no se regeneran.
* En mitocondrias y cloroplastos los centros de obtención de energía se sitúan en las membranas, al igual que ocurre en las bacterias. Por otro lado, los tilacoides que encontramos en cloroplastos son similares a unos sistemas elaborados de endomembranas presentes en cianobacterias.
* En general, la síntesis proteica en mitocondrias y cloroplastos es autónoma.
* Algunas proteínas codificadas en el núcleo se transportan al orgánulo, y las mitocondrias y cloroplastos tienen genomas pequeños en comparación con los de las bacterias. Esto es consistente con la idea de una dependencia creciente hacia el anfitrión eucariótico después de la endosimbiosis. La mayoría de los genes en los genomas de los orgánulos se han perdido o se han movido al núcleo. Es por ello que transcurridos tantos años, hospedador y huésped no podrían vivir por separado.
* En mitocondrias y cloroplastos encontramos ribosomas 70s, característicos de procariotas, mientras que en el resto de la célula eucariota los ribosomas son 80s.
* El análisis del RNAr 16s de la subunidad pequeña del ribosoma de mitocondrias y plastos revela escasas diferencias evolutivas con algunos procariotas.
* Una posible endosimbiosis secundaria (es decir, implicando plastos eucariotas) ha sido observado por Okamoto e Inouye (2005). El protista heterótrofo Hatena se comporta como un depredador e ingiere algas verdes, que pierden sus flagelos y citoesqueleto, mientras que el protista, ahora un anfitrión, adquiere nutrición fotosintética, fototaxia y pierde su aparato de alimentación.
* Ni las mitocondrias ni los plastos pueden sobrevivir fuera de la célula. Sin embargo, este hecho se puede justificar por el gran número de años que han transcurrido: los genes y los sistemas que ya no eran necesarios fueron suprimidos; parte del ADN de los orgánulos fue transferido al genoma del anfitrión, permitiendo además que la célula hospedadora regule la actividad mitocondrial.
* La célula tampoco puede sobrevivir sin sus orgánulos: esto se debe a que a lo largo de la evolución gracias a la mayor energía y carbono orgánico disponible, las células han desarrollado metabolismos que no podrían sustentarse solamente con las formas anteriores de síntesis y asimilación.
MORALEJA
2.100 MILLONES DE AÑOS
FÓSILES DE GUNFLINT
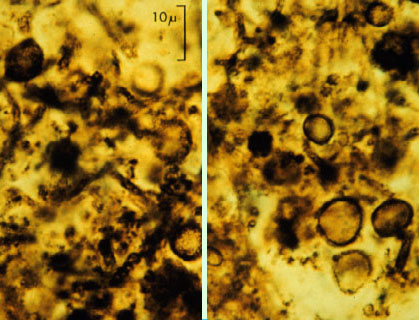
Los fósiles de Gunflint (Australia) son organismos capaces de metabolizar O2..También hace unos 2.100 Ma aparecen las primeras células Eucariotas (PROTISTAS). Son tal vez parecidas a algas verdes fotosintéticas unicelulares, y han recibido el nombre de Grypania.
El Reino Protista está conformado por un grupo de organismos que presentaban un conjunto de características que impiden colocarlos en los reinos ya existentes de una manera plenamente definida. Esto se debe a que algunos protistas pueden parecerse y actuar como individuos del reino plantas, otros protistas pueden parecerse y actuar como organismos del reino animal, pero los organismos del reino protista no son ni animales ni plantas.
Los individuos del reino de los protistas son los que presentan las estructuras biológicas más sencillas entre los eucariotas (ya que su ADN está incluido en el núcleo de la célula), y pueden presentar una estructura unicelular (siendo esta la más común), multicelular o colonial (pero sin llegar a formar tejidos). Los protistas son autótrofos (en su mayoría) y producen un alto porcentaje del oxígeno de la tierra. Es decir,son una célula intermedia entre las procariotas y eucariotas.

Grypania spiralis
Grypania spiralis es un fósil del Paleoprotozoico (hace unos 2.100 millones de años). Pudo ser una colonia gigante de bacterias, pero, dado su tamaño (más de un centímetro), se cree que pudo ser una alga filamentosa, es decir, un organismo eucariota; de ser así, sería el primer miembro conocido del dominio Eukarya Los fósiles fueron hallados en la formación Negaunee Iron en Palmer (Michigan).
2.023 MILLONES DE AÑOS
PERIODO OROSIDICO

El período Orosírico (2.050 – 1.800 Ma), también llamado Orosiriense, es la tercera de las subdivisiones de la era Paleoprotozoica; está caracterizado por los impactos de dos grandes asteroides, uno marcando su comienzo (hace 2.023 Ma) y otro su final (hace 1.850 Ma), por el inicio de una nueva etapa de rift oceánico y por la intensa etapa orogénica que tiene lugar en todos los continentes (de ahí el nombre del período, “orosira”, que significa “cordillera”).
A comienzos del período Orosírico, hace 2.023 Ma, se produjo el impacto de un gran asteroide (de entre 5 y 10 km de diámetro) sobre la superficie terrestre, en Sudáfrica, dando como resultado el conocido cráter de Vredefort (el más grande del mundo con 300 km de extensión). Pese a la espectacularidad del impacto, el acontecimiento no perjudicó de manera notable el desarrollo de los seres vivos que vivían en la Tierra.

Cráter de Vredefort
La elevación central del cráter se conoce como domo de Vredefort. Se calcula que al impactar el meteorito se liberó una energía de 100 millones de megatones de dinamita, y que se desplazaron 70.000 km³ de roca.
La proliferación de organismos fotosintéticos oxigénicos que habían aparecido hasta entonces, habían enriquecido la atmósfera con oxigeno y los cielos, que hasta entonces habían sido rojizos,fueron tomando el color azul que conocemos ahora. La atmósfera había sido reductora , a comienzos de este periodo se convirtió en medioxidante.
Entre 2000 y 1900 Ma , la corteza terrestre sufrió numerosas orogenias en todos los continentes,como por ejemplo, la orogénia de Glenburgh(2005-1920 Ma),las orogénias Hudson,Kareliense,Ruker y Svecofenniense (2000 Ma),la orogénia Churchiliana (1900 Ma) y la segunda fase de la orogenia Nagssugtoqidiense(1900 Ma) que se produjeron a la par de la orogenia Ebúrnica,que ya había comenzado hace 2200 Ma en algunas regiones.
Este conjunto de orogenias provocaron, hace unos 1.900 Ma, un nuevo cambio en el funcionamiento de la corteza, ganando importancia los procesos de rift oceánico por encima de la acreción vertical, por lo que se considera que a partir de ese momento acabó la denominada fase de transición de la evolución cortical y empezó la fase de la tectónica de placas (con un comportamiento de la corteza muy similar al que hay en la actualidad). Los continentes, que ocupaban un 53% de la superficie actual, disponían ya de unos cratones lo suficientemente sólidos como para soportar las cadenas generadoras de flysches (sucesiones potentes de materiales marinos profundos) y molasas (sedimentos marinos someros y no marinos originados por erosión de cinturones montañosos tras su levantamiento por efectos de una orogenia).
A estas alturas el volumen de los océanos era muy similar al que tenemos hoy en día.
Hace unos 1.870 – 1.800 Ma se inició una segunda etapa orogénica que duraría varias decenas de millones de años, con orogenias destacables, como las orogenias Barramundi (1.870 – 1.800 Ma), Kimbiense (1.845 – 1.700 Ma) y Penokiense (1.850 – 1.840 Ma).
Hace 1.850 Ma, coincidiendo con el final de la orogenia Churchilliana, la Tierra sufrió el impacto de otro gran asteroide (de unos 10-15 km de diámetro), que dio lugar al actual cráter de Sudbury (localizado en Ontario, Canadá), el segundo cráter descubierto más grande del mundo.
Las orogenias Hudson y Barramundi finalizaron hace unos 1.800 Ma, poniéndole punto y final a este período.
MORALEJA
Definitivamente la tierra y la vida que hoy conocemos ,se produjo gracias a los miles de impactos que fue sufriendo el planeta.
1.800 MILLONES DE AÑOS
PERIODO ESTATÉRICO
El período Estatérico o Estateriense (del griego : statheros, que significa «estable, firme»), una división de la escala temporal geológica, es el cuarto período geológico de la Era Paleoprotozoica. Comienza hace 1.800 millones de años y finaliza hace 1.600 millones.
El período se caracteriza en la mayoría de los continentes por la formación de nuevas plataformas o por la cratonización (masas de tierra flotantes) final de los cinturones de plegamiento. El supercontinente Columbia se formó a comienzos de este período.
Durante este período probablemente apareció la primera célula eucariota.
MORALEJA
Las grandes masas de tierra se ha ido desplazando y chocando unas con otras desde el inicio de nuestro planeta,de hecho aun siguen en movimiento.Por lo tanto la tierra aun sigue creandose a si misma y evolucionando.
NENA
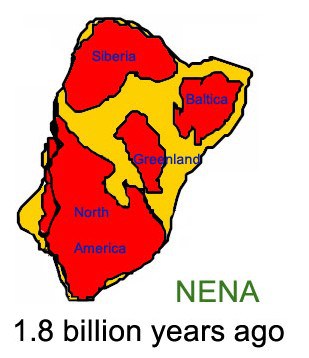
Nena fue un supercontinente que surgió hace aproximadamente 1.800 millones de años y que comprendía Siberia, Báltica, Groenlandia y Norteamérica. Nena se unió a Atlántica y Ur (que ya era más extenso) para luego formar Rodinia. El término Nena es un acrónimo que deriva de «Norte de Europa y Norte de América».
COLUMBIA Y ATLÁNTICA
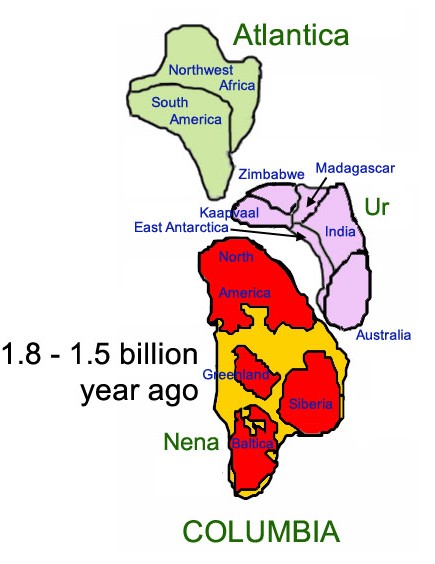
Columbia (también conocido como Nuna y, más recientemente, Hudsonlandia o Hudsonia) es el nombre de uno de los supercontinentes postulados de la tierra. Existió hace aproximadamente de 1800 a 1500 millones años en el Paleoproterozoico, siendo el supercontinente más antiguo.Consistió en un proto-cratón que integraban los ex-continentes de Laurentia, Báltica, Ucrania, Amazonia, Australia, y posiblemente Siberia, norte de China y Kalahari. La existencia de Columbia se basa en datos paleomagnéticos.
Se estima que Columbia tendría cerca de 12.900 kilómetros de norte a sur, y cerca de 4.800 kilómetros en su parte más ancha. La costa del este de la India estaba unida a Norteamérica occidental, con Australia meridional y Canadá occidental. La mayor parte de América del Sur estaba girada de manera que el borde occidental (lo que hoy en día es Brasil) se alineó con el este de América del Norte, formando un margen continental que se extendía hasta el sur de Escandinavia
Atlántica fue un supercontinente que surgió hace aproximadamente 1.800 millones de años. Alrededor de 200 millones de años más tarde, se convirtió en parte del supercontinente Columbia y 300 millones de años más tarde, se separa de Columbia. Hace 1.100 millones de años (o 400 millones de años después de la desintegración de Columbia), se une a Nena y Ur pasa a formar parte del supercontinente Rodinia. Después de que Rodinia se dividiera y los fragmentos se volvieran a reunir hace 600 millones de años, pasa a formar parte del supercontinente Pannotia.
1.600 MILLONES DE AÑOS
ERA MESOPROTEROZOICA
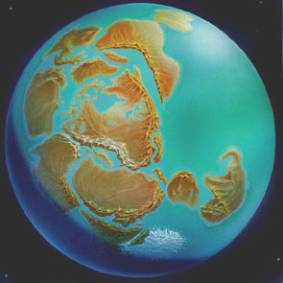
Rodinia
La era Mesoproterozoica (1.600 – 1.000 Ma) es la segunda de las subdivisiones del eón Proterozoico, y está caracterizada por ser la época de máxima diversidad y abundancia de los estromatolitos y por la ruptura del supercontinente Columbia y la formación del supercontinente Rodinia. En el Mesoproterozoico también empiezan a aparecer fósiles (tales como Bangiomorpha pubescens) que se parecen a ciertas algas rojas actuales (aunque resultan dudosos, pues podrían ser colonias de cianobacterias), y fósiles que quizá fueran los predecesores del linaje que condujo a los hongos (aunque también podrían ser colonias de actinobacterias filamentosas).
PERIODO CALÍMICO
El período Calímico (1.600 – 1.400 Ma), del griego calymma, que significa “cubierta”, es la primera de las subdivisiones de la era Mesoproterozoica; está caracterizado por una intensa expansión de los depósitos sedimentarios sobre las plataformas continentales desarrolladas a lo largo y ancho del supercontinente Columbia y por la formación de nuevas plataformas, todo ello coincidiendo con las últimas etapas de la segunda fase de la orogenia Nagssugtoqidiense (iniciada hace 1.900 Ma). Esta orogenia finalizó hace unos 1.500 Ma, momento en que empezó la fragmentación del supercontinente Columbia y comenzaron los depósitos de grandes cantidades de materiales volcanosedimentarios.
1.500 MILLONES DE AÑOS
CÉLULAS EUCARIOTAS
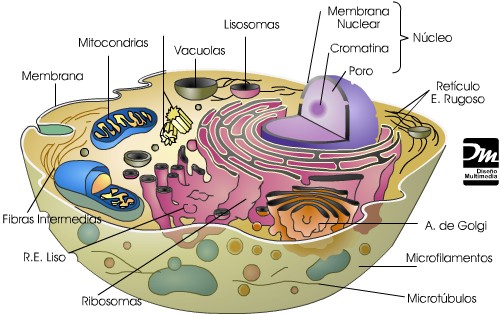
Célula eucariota animal
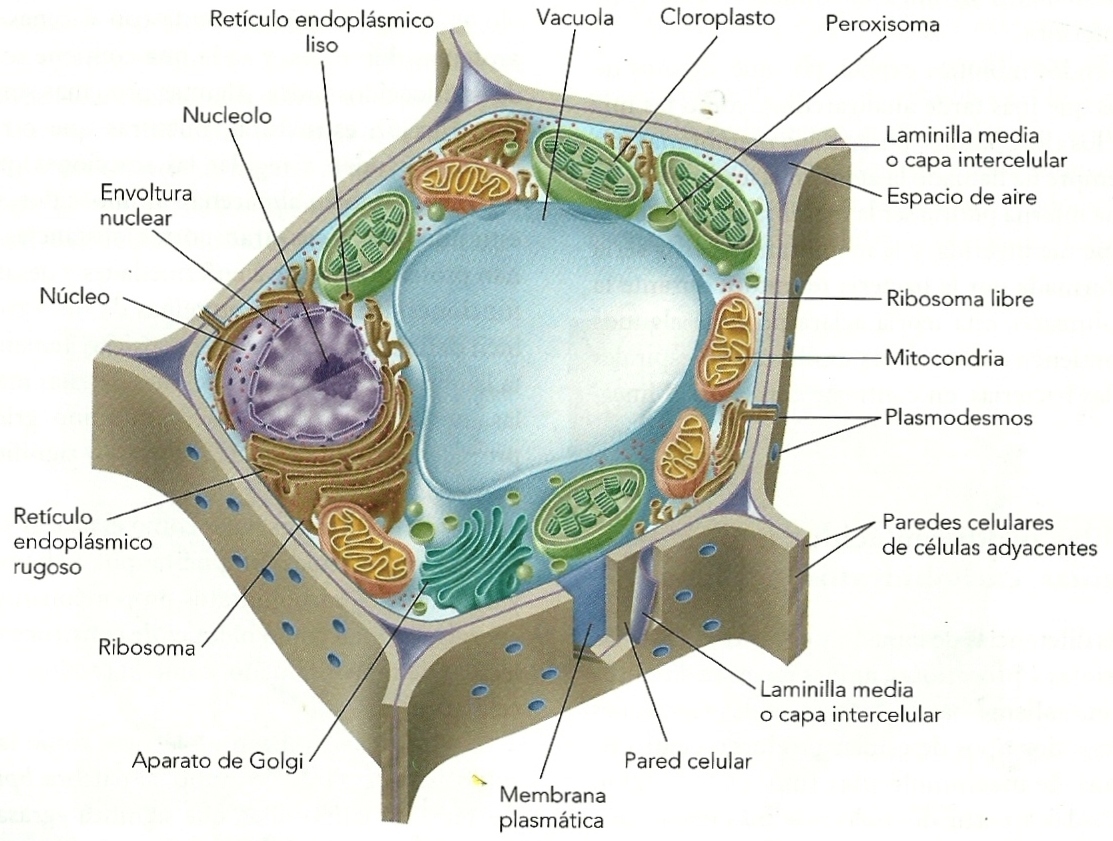
Célula eucariota vegetal
Las células eucariotas tienen su información genética encerrada dentro de la envoltura nuclear. Su citoplasma presenta orgánulos interconectados cuyos límites se encuentran fijados por membranas biológicas. El compartimiento más notorio del protoplasma es el núcleo.
Las eucariotas suelen contener mitocondrias, que son orgánulos membranosos que producen energía. Algunas eucariotas protistas, sin embargo, ya no exhiben mitocondrias tras el curso de la evolución. La presencia de plastos en el citoplasma, por otra parte, permite que ciertas eucariotas puedan realizar la fotosíntesis.
Pese a la variedad de eucariotas, estas células comparten una misma composición bioquímica y un metabolismo homogéneo, hecho que representa una importante diferencia respecto a las procariotas, las células cuyo material genético se encuentra repartido en diversos organelos.
Cabe destacar que los organismos eucariontes constituyen el dominio Eukarya, que incluye seres de los cuatro reinos: animales, plantas, hongos y protistas.
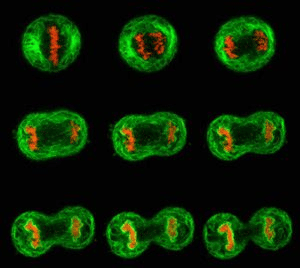
Con respecto a su reproducción, los eucarionte son capaces de dividirse de manera asexual (fenómeno conocido como mitosis), aunque en general atraviesan procesos reproductivos de tipo sexual que se basan en la meiosis y que no se ve en las células procariotas.
La Meiosis es una de las formas de la reproducción celular. Este proceso se realiza en las glándulas sexuales para la producción de gametos. Es un proceso de división celular en el cual una célula diploide experimenta dos divisiones sucesivas, con la capacidad de generar cuatro células haploides . En los organismos con reproducción sexual tiene importancia ya que es el mecanismo por el que se producen los óvulos y espermatozoides (gametos).Este proceso se lleva a cabo en dos divisiones nucleares y citoplasmáticas, llamadas primera y segunda división meiótica o simplemente meiosis I y meiosis II. Ambas comprenden profase, metafase, anafase y telofase.

En la interfase se duplica el material genético. En meiosis I los cromosomas homólogos se reparten en dos células hijas, se produce el fenómeno de entrecruzamiento. En meiosis II, al igual que en una mitosis, cada cromátida migra hacia un polo. El resultado son 4 células hijas haploides .
Durante la meiosis I miembros de cada par homólogo de cromosomas se emparejan durante la profase, formando bivalentes. Durante esta fase se forma una estructura proteica denominada complejo sinaptonémico, permitiendo que se produzca la recombinación entre ambos cromosomas homólogos. Posteriormente se produce una gran condensación cromosómica y los bivalentes se sitúan en la placa ecuatorial durante la primera metafase, dando lugar a la migración de cromosomas a cada uno de los polos durante la primera anafase. Esta división reduccional es la responsable del mantenimiento del número cromosómico característico de cada especie. En la meiosis II, las cromátidas hermanas que forman cada cromosoma se separan y se distribuyen entre los núcleos de las células hijas. Entre estas dos etapas sucesivas no existe la etapa S (replicación del ADN). La maduración de las células hijas dará lugar a los gametos.
Para reproducirse, las eucariotas alternan las generaciones haploides, donde se encuentra tan sólo un ejemplar de cada cromosoma, y las diploides, que tienen el doble.
Para pasar del primero al segundo tipo de generación es necesario atravesar la fusión nuclear, o fecundación, y volver al primero mediante la meiosis. Esto no quiere decir, por otro lado, que no existan ciertas diferencias importantes entre los eucariontes. Cabe mencionar que, en comparación con los procariontes, sus tasas de metabolismo son menores y el tiempo que necesitan para la generación es mayor, y su superficie no es tan representativa de su volumen.
Continuando con el proceso reproductivo, cabe mencionar que puede tener lugar de tres formas bien diferenciadas:
* la bipartición: consiste en la división de una célula en otras dos que sean absolutamente iguales;
* la gemación: cuando se forma una especie de tumor en una célula, el cual crece durante un tiempo hasta convertirse en una célula independiente;
* la esporulación: una célula comienza por dividir su propio núcleo en copias más pequeñas, y finalmente hace lo mismo con su citoplasma, para que se formen más células.
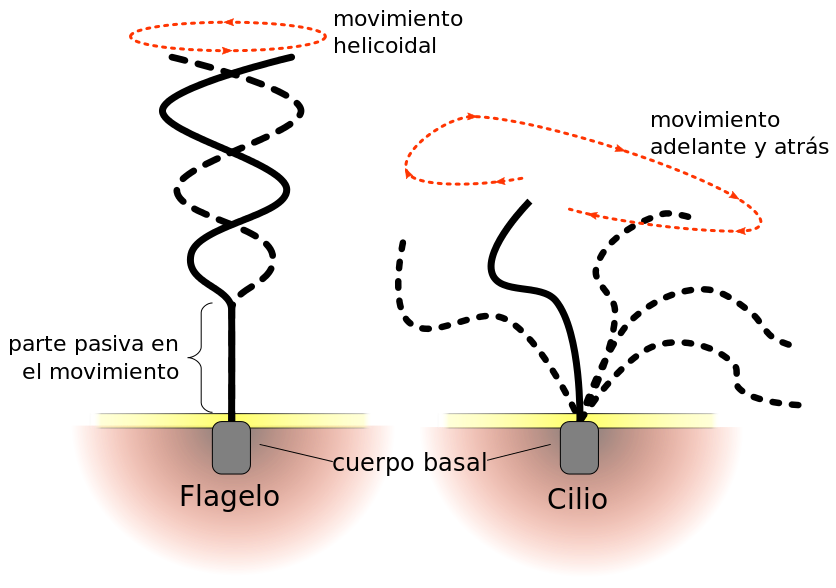
En las células eucariotas se encuentran uno o dos ejemplares de una estructura conocida como flagelo. Es posible distinguir dos patrones de relación entre las células y sus flagelos: las acrocontas se trasladan detrás de su o sus flagelos, mientras que las opistocontas lo hacen por detrás. Esto último se da en el reino de los hongos y el de los animales, y es posible advertir dicho comportamiento en un espermatozoide de nuestra especie.
Los flagelos eucariotas pertenecen a una agrupación de estructuras que se conoce con el nombre de undulipodios, donde también se cuentan los cilios. Una diferencia básica entre ambos es que los flagelos cobran un mayor tamaño, dado que se complican con otras estructuras añadidas. Básicamente, tanto los flagelos como los cilios presentan un aspecto cilíndrico y parejo a lo largo de todo su cuerpo y con sus extremos redondeados. Su núcleo, que tiene una estructura llamada anoxema, está recubierto por un tejido plasmático, lo cual permite que el citoplasma de la célula lo penetre y acceda a su interior.
MORALEJA
Las células eucariotas fueron un gran paso para la vida ya que de ellas estamos compuestos todos los seres vivos del planeta.No cabe mencionar su parecido con la imagen de un espermatozoide que todos conocemos.
1.400 MILLONES DE AÑOS
PERIODO ECTÁSICO

Isla Somerset
El período Ectásico (del griego «ectasis» que significa «extensión»), una división de la escala temporal geológica, es el segundo período geológico de la Era Mesoproterozoica. Comienza hace 1.400 millones de años y finaliza hace 1.200 millones de años.
El nombre de este período se justifica por la expansión continua de los depósitos sedimentarios y volcánicos sobre las plataformas continentales durante este período.
Se han identificado fósiles de Bagiomorpha pubescens en rocas de hace 1.200 millones de años de la formación Hunting de la isla Somerset, Canadá. Podrían corresponder a un alga roja, y por tanto, al organismo pluricelular y con reproducción sexual más antiguo conocido hasta ahora.Sin embargo, resultan dudosos y podrían ser realmente colonias de cianobacterias.
1.200 MILLONES DE AÑOS
PERIODO ESTÉNICO
El período Esténico (del griego -stenos- que significa «estrecho»), una división de la escala temporal geológica, es el tercer período geológico de la Era Mesoproterozoica. Comienza hace 1200 millones de años y finaliza hace 1100 millones de años.El nombre se justifica en los estrechos cinturones polimetamórficos formado durante este período. El supercontinente Rodinia se formó en el Esténico.
Hace unos 1200 millones de años, en el límite Ectásico-Esténico, se produjo uno de los mayores episodios magmáticos de la de la tierra, debido probablemente a una gigantesca avalancha mantélica (son desplomes hipotéticos del manto terrestre. Se piensa que la avalanchas mantélicas se originan a 660 km de profundidad, lugar en el que partes frías del manto que se encuentran descendiendo se apilan para luego descender esporádicamente cuando se ha acumulado cierta cantidad crítica de material). Otros episodios similares se han producido aproximadamente cada 800 millones de años: el primero y más intenso hace 2700 Ma, en el Neoarcaico, y otro, de menor intensidad que el anterior pero mayor que este, hace 1900 Ma, en el Orosirico
1.100 MILLONES DE AÑOS
RODINIA
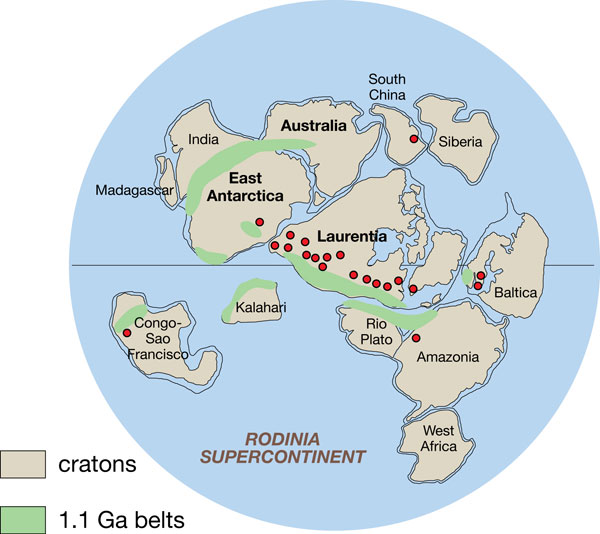
Rodinia (del ruso , ródina, patria) fue un supercontinente que existió hace 1100 millones de años, durante la Era Neoproterozoica. estaba compuesta de la mayoría de la tierra emergida hasta ahora. Empezó a fracturarse hace 800 millones de años debido a movimientos magmáticos en la corteza terrestre, acompañados por una fuerte actividad volcánica. La existencia de Rodinia se basa en pruebas de paleomagnetismo que permiten obtener la paleolatitud de los fragmentos, pero no su longitud, que los geólogos han determinado mediante la comparación de estratos similares, actualmente muy dispersos.
Rodinia se formó y se deshizo durante el Neoproterozoico. Probablemente existió como un único continente, hasta que comenzó a fragmentarse en ocho pequeños continentes hace alrededor de 800 millones de años. Se cree que la acumulación de tierra en un solo lado del planeta fue responsable en gran parte del clima frío del Neoproterozoico. Rodinia comenzó a formarse hace alrededor de 1.300 millones de años a partir de tres o cuatro continentes preexistentes, un acontecimiento conocido como la orogenia Grenville.
Rodinia probablemente estuvo centrada al sur del ecuador.Durante el periodo criogénico (850 Ma), gran parte de Rodinia pudo haber estado cubierta por glaciares o formando parte del casquete de hielo del Polo Sur. El interior del continente, distante de los efectos moderadores del océano, es probable que fuera estacionalmente muy frío. Rodinia estaba rodeado por el superocéano que los geólogos denominan Mirovia (de Mir, la palabra rusa que significa «paz»).
Las temperaturas frías puede que fueran acentuadas durante las primeras etapas de la dislocación continental. Los picos de calentamiento geotérmico dislocarían la corteza y las rocas se elevarían en relación con su entorno. Esto crearía zonas de mayor altitud, donde el aire es más frío y es menos probable que el hielo se funda con los cambios estacionales, y puede explicar la evidencia de abundante glaciación durante el Período Ediacárico. La separación final de los continentes creando nuevos océanos y expandiendo el fondo oceánico, con producción de rocas menos densas, probablemente incrementó el nivel del mar por desplazamiento del agua de los océanos. El resultado fue un mayor número de océanos de aguas someras.
La evaporación del agua de los océanos pudo hacer que aumentaran las precipitaciones, lo que a su vez aumentaría la erosión de la roca expuesta. Si en los modelos de simulación por computador se introduce la relación de isótopos estables de 18O:16O, se comprueba que, además de la fuerte erosión de la roca volcánica, este aumento de las precipitaciones hizo reducir los niveles de los gases de efecto invernadero por debajo del umbral que activaría el período de glaciación extrema conocida como la «Tierra bola de nieve». Toda esta actividad tectónica introduciría además en el medio marino nutrientes biológicamente importantes, lo que pudo haber desempeñado un papel importante en el desarrollo de los primeros animales.
Rodinia comenzó a fragmentarse a más tardar hace 750 millones de años.Otros continentes, incluyendo Báltica y Amazonia, se dislocaron de Laurentia hace 600-550 millones de años, abriendo el océano lapetus entre ellos. La separación también llevó al nacimiento de océano Pantalassa (o Paleo-Pacífico).Los ocho continentes que formaban parte de Rodinia más tarde volvieron a reunirse en el supercontinente global denominado Pannotia, y después una vez más como Pangea.
1.000 MILLONES DE AÑOS
ERA NEOPROTEOZOICA
El Neoproterozoico, una división de la escala temporal geológica, fue la última era geológica de las tres que componen el Eón Proterozoico; comenzó hace 1000 millones de años y terminó hace 542 Ma. La nomenclatura para denominar a este período ha sido un tanto inestable.
A finales de esta era Rodinia comenzó a fragmentarse en ocho continentes.Durante el periodo criogénico que se comentara mas adelante se producen dos episodios glaciares importantes dando la lugar a la primera extinción en masa documentada en el registro fósil.estos ocho continentes se volvieron a unir dando lugar a Pannotia.
La idea de como fue el neoproterozoico es relativamente reciente(1960). las células eucariotas dieron un paso mas,dando lugar a la presencia de acritarcos (cualquier pequeña estructura orgánica (ni carbonatada ni silicia) no soluble en ácido, que no pueda ser clasificada de otra manera),así como microfósiles mineralizados tecamebinos (amebas con caparazón orgánico o mineralizados),tintínidos (son formas unicelulares, relativamente grandes, con una estructura interna compleja) o foraminíferos,siendo los primeros protistas heterótrofos planctónicos que vivían como micropredadores en las costas en esta era.Posiblemente se produjo la radiación de las algas Rhodophyta y las Chiorophyta.

Fósil ediacárico de Cyclomedusa
Los Protistas dieron origen a los los primeros metazoos, animales invertebrados conocidos como la Fauna de Ediacara por haberse encontrado sus fósiles en las colinas de Ediacara, una región del sur de Australia. Estos primeros animales pluricelulares tenían cuerpo blando y aplastado y muchos tenían formas variadas y extrañas: algunos tienen un cuerpo en forma de disco, otros parecen una cinta larga; los más destacados son Spprigina, Tibrachidium, los primeros gusanos,medusas,artrópodos etc., que dominaron los océanos evolucionando y expandiéndose. Así pues, excepto metazoos y plantas de los que no tenemos registro el resto de los grupos de organismos ya estaban presentes en el Tónico.
PERIODO TÓNICO
El período Tónico (del griego , tonas, que significa «estiramiento»), es una división de la escala temporal geológica, es el primer período geológico de la Era Neoproterozoica. Comienza hace 1.000 millones de años y finaliza hace 850 millones de años.
En este periodo comenzó la disolución de Rodinia y aparecieron los acritarcos.
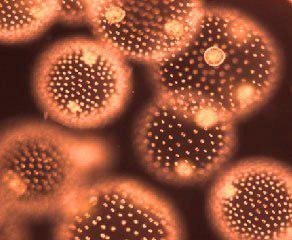
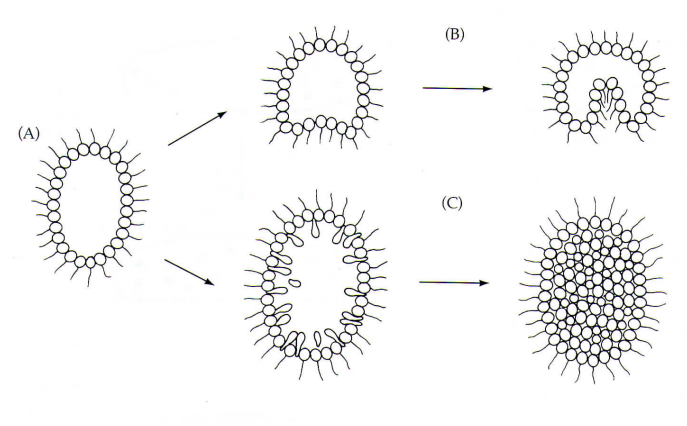

Hongos mucilaginosos
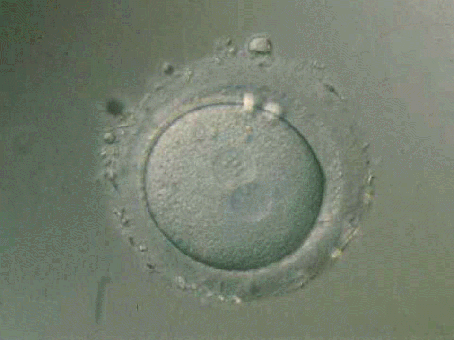
Cigoto
MORALEJA
Esto es el comienzo de la evolución humana el mas grande se come a el mas pequeño.Solo hay que mirar un poco atrás para ver que las cosas no han cambiado,es la ley del mas fuerte.
950 MILLONES DE AÑOS
GLACIACIÓN STUARTIANA-VARANGIANA
Poco se puede decir de esta época mas que fue el comienzo de una de las glaciaciones mas importantes que ha sufrido el planeta.dando lugar aun planeta yermo y frió durante millones de años.
850 MILLONES DE AÑOS
PERIODO CRIOGÉNICO

El período Criogénico (del Griego cryos, «hielo» y génesis, «origen»), una división de la escala temporal geológica, es el segundo período geológico de la Era Neoproterozoica. Comienza hace 850 millones de años y finaliza hace 630. Su nombre hace referencia a los depósitos glaciales encontrados en latitudes tropicales, seguidos de sedimentos de carbonatos . Los glaciares se extendieron y retrocedieron en una serie de pulsos rítmicos, posiblemente alcanzando el ecuador.
hay una hipótesis llamada «Tierra bola de nieve» que postula que esta glaciación fue tan grande que se congelaron hasta los océanos.
Durante el Criogénico, el supercontinente Rodinia se fragmentó y el supercontinente Pannotia comenzó a formarse.Se cree que por este motivo comenzó la glaciación ,la ruptura de Rodinia expone a la materia orgánica a la respiración anaerobia,creando una atmósfera en la que el gas de efecto invernadero es metano.El metano atmosférico reacciona con el oxigeno en contraste con el CO2 .Esto produjo que el metano disminuyera drasticamente y enfriase la tierra.estas temperatura bajas impedían crear mas metano por lo tanto la tierra se enfrió aun mas,llegando a congelar los océanos.
TIERRA BOLA DE NIEVE

La teoría de la Tierra bola de nieve (1998)(Paul Hoffman y Daniel Schrag) se basa en un descenso inimaginable de la temperatura del planeta cubriendo de hielo la tierra incluyendo los mares.la temperatura media debió ser de -50ºC ya que la radiación solar era reflejada hacia la superficie por el hielo.Los océanos estaban congelados ,por lo tanto no podían absorber calor,haciendo que la temperatura variase notablemente entre el día y la noche.
El motivo de este descenso tan drástico de temperatura pudo deberse a la reducción del efecto invernadero que incrementa la temperatura atmosférica por concentraciones de dióxido de carbono,vapor de agua y metano que absorben y retienen la radiación térmica .Si estos gases no la contuviesen ,esta se escaparía hacia el espacio.Al disminuir estos gases provocarían el enfriamiento de la tierra.
Es decir,los volcanes emiten dióxido de carbono a la atmósfera,mientras que la meteorización de rocas silicatadas ,en las que intervienen la acción del agua y la actividad de las plantas (en este caso algas),lo retira de la atmósfera absorviendolo en las rocas.los organismos vivos extraen este dióxido de carbono y es rápidamente devuelto a la atmósfera.Este proceso se produce con mayor intensidad en las zonas mas cálidas ,ralentizando el proceso.Durante el criogénico los continentes estaban situados en los trópicos donde hacia mas calor y por lo tanto mas consumo de dióxido de carbono haciendo que la carencia de este en la atmósfera ,disminuyera el efecto invernadero.
El planeta entro en un periodo glacial tan frió que exterminó a la mayoría de los seres vivos del planeta ,y los poco que sobrevivieron se limitaron a subsistir en un clima tan frió.
Se cree que gracias a las cianobacterias y a su producción de oxigeno ,se fueron mutando hasta conseguir cianobacterias de bióxido de carbono.Esta acumulación de dióxido de carbono pudo salvar al planeta de esta glaciación.
MORALEJA
Es solo una teoría ,en la que todos no están de acuerdo.Yo tampoco creo que si se hubiera congelado por completo la vida se habría abierto camino.Ya que la energía solar es necesaria para la fotosíntesis y la proliferación de cianobacterias.Supongo que si se congelo en su mayor parte ,aun prevaleciendo zonas cálidas que ayudarían al proceso de descongelación.
635 MILLONES DE AÑOS
PERIODO EDIACÁRICO

El período Ediacárico (nombrado por la montañas Ediacara), una división de la escala temporal geológica, es el tercer y último período geológico de la Era Neoproterozoica. Comienza hace unos 635 millones de años y finaliza hace 542 millones de años.Durante este periodo los fragmentos de Rodinia se vuelven a unir para formar Pannotia que acabara por fragmentarse a finales e este periodo.
Este período es quizás el origen de los animales, hace unos 570 millones de años y es famoso por su «fauna ediacárica». Son los fósiles de organismos pluricelulares con diferenciación de tejidos más antiguos conocidos que no resultan dudosos. Todavía no habían desarrollado caparazones o esqueletos y su relación con los organismos actuales o incluso con los posteriores de la explosión cámbrica es difícil de interpretar. Los tipos más comunes se parecen a gusanos segmentados, frondas, discos o bolsas inmóviles. Hay científicos que creen que estos fósiles representan invertebrados primitivos que fueron ancestros de los animales modernos, u organismos que no son animales verdaderos. Dado que los fósiles no muestran mucho sobre su posición en la línea evolutiva, es posible que su origen e historia jamás se descubran.
600 MILLONES DE AÑOS
PANNOTIA
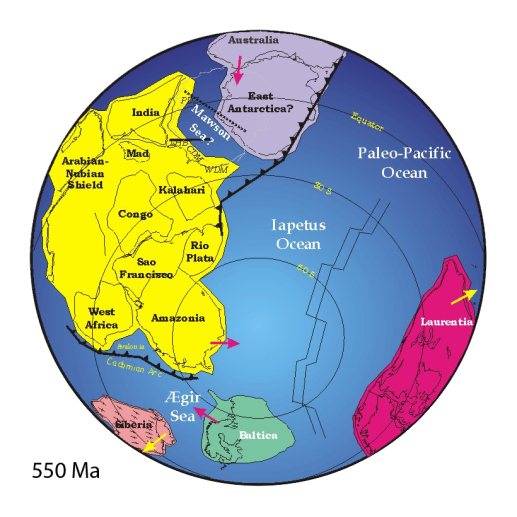
Pannotia fue un supercontiente de corta duración; se cree que no alcanzó a terminar de formarse cuando empezaron los esfuerzos distensivos, magmatismo de intraplaca y demás hechos geológicos que hicieron que se disgregara. Los fragmentos que lo formaron y que luego se disgregaron existen hasta el día de hoy; son los cratones de Australia, Antártida del Este, Madagascar, Laurentia, Báltica, Siberia, Amazonia, África del Oeste, Río de la Plata, San Francisco, Congo, India, Kalahari, y Arábico-Nubaniano. Todos ellos al generarse Pannotia y posteriormente a fracturarse, se unieron entre sí hasta la configuración actual continental; de la separación de Pannotia quedaron 4 grandes porciones de tierra: Laurentia, Gondwana (contenía los cratones de Antártida, India, Amazonia, África del oeste, Río de la Plata, San Francisco entre otros), Báltica y Siberia.
570 MILLONES DE AÑOS
FAUNA DE EDIACARA

La fauna de Ediacara también conocida como biota de ediacara o biota vendiense son antiguas formas de vida sésiles (sin un órgano que sirva de pie o soporte)con forma tubular (de tubo) y de hoja que habitaron en el planeta durante el periodo ediacárico y son los organismos pluricelulares complejos mas antiguos.
La aparición y desaparición de estos organismos fue breve coincidiendo con la explosión cámbrica,siendo sustituidos por organismos nuevos.La mayoría de los fósiles de esta época no se asemejan a la forma vida posterior del planeta,por lo tanto, algunos paleontologos los han clasificado como una rama independiente e extinta,denominada Vendozoa o Vendobionta. Otros simplemente creen que fue un experimento fallido de la futura evolución de los seres vivos.
Los primeros fósiles fueron descubiertos por Alexander Murray (geologo) pero aun no tenian muy claro si eran fosiles y no se le dió mucha importancia.Otros geologos de los años 30 también hicieron sus descubrimientos pero tampoco se les dió importancia pues no tenian una referencia anterior con que compararlos ,pensaron que eran erosiones de la tierra.
Los fósiles de Ediacara eran tan extraños e inesperados que su descubridor, un joven geólogo australiano llamado Reginald Sprigg, se pasó años presentándolos a congresos y mandando sus resultados a revistas, sin que nadie le creyera. Estaba haciendo un estudio sobre unas minas abandonadas en el sur de Australia, y un día, cuando estaba almorzando, volvió distraídamente una piedra arenisca y se quedó anonadado. Aquellas rocas eran del principio del Cámbrico, de hace casi 600 millones de años, y hasta entonces sólo se conocían fósiles del Cámbrico avanzado (de hace 540 a 520 millones de años). Pero los que veía no se parecían en nada a los que se habían encontrado: eran huellas delicadas, casi evanescentes, de pequeños organismos de formas diversas, normalmente de disco, cinta o pluma. Algunos de ellos parecen arrastrar suaves flecos. Nadie había visto nunca nada similar, aunque algunos de estos seres recordaban a medusas o gusanos.
No fue hasta 1957 con el descubrimiento de Charnia que se empezara a tomar en serio el tema. Este fósil en forma de fronda fue descubierto en el bosque de Charnwood, en Inglaterra, y a causa de los detallados mapas geológicos del British Geological Survey no cupo ninguna duda que estos fósiles estaban insertos en rocas precámbricas. El paleontólogo Martin Glaessner trazó finalmente la conexión entre este descubrimiento y los anteriores, y con una combinación de dataciones más precisas de los ejemplares existentes y una inyección de vigor a la investigación, se descubrieron muchos más miembros de la fauna ediacárica.
Sin embargo,hasta 1967 los fósiles no se distiguian muy bien ya que estaban incrustados en arenisca pero con el descubrimiento de S.B.Misra de fósiles en cenizas fue mas fácil interpretarlos.
La causa de su fosilización no esta muy clara,aunque la hipótesis mas extendida es que fueron rápidamente cubiertos de ceniza o de arena ,atrapándolos junto al barro o los tapetes de microbios en los que vivían.
Su tamaño podía oscilar entre pocos milímetros hasta varios metros,podían ser simples manchas en la roca hasta figuras mas complicadas.Se diferenciaban de los organismos ya conocidos en su variedad de tipos.Estos organismos podían dividirse según la siguiente variedad morfológica:
Embriones: Los descubrimientos recientes de vida pluricelular precámbrica documentan numerosos casos de fósiles con forma de embriones, especialmente en la Fm. Doushantuo, en China. Algunos hallazgos despertaron un gran interés mediático aunque algunos investigadores han afirmado que, en realidad, son estructuras no orgánicas formadas por la precipitación de minerales dentro de un hueco. Otros embriones han sido interpretados como restos de bacterias reductoras de azufre gigantes similares a Thiomargarita.
Recientemente se ha visto, mediante el análisis de muestras de estos especímenes por microscopía tomográfica de rayos X de fuente sincrotrón, que no se trata de embriones ni de ninguna otra otra estructura asociada a los metazoos, sino que más bien se trata de acumulaciones de células semejantes a quistes producto de una división celular de tipo politómico, es decir, que proceden de una célula progenitora que aumenta mucho de tamaño, produciéndose posteriormente divisiones celulares rápidas sucesivas con poco o ningún crecimiento entre etapas.
 Discos:Fósiles con forma circular, como Ediacaria, Cyclomedusa y Rugoconite, hicieron que los fósiles ediacáricos fueran identificados originalmente como cnidarios, un grupo que también incluye las medusas y los corales. Debido a que sólo el dorso inferior del organismo se ha fosilizado, no se conocen las suficientes características útiles del organismo para establecer una estructura adecuada.
Discos:Fósiles con forma circular, como Ediacaria, Cyclomedusa y Rugoconite, hicieron que los fósiles ediacáricos fueran identificados originalmente como cnidarios, un grupo que también incluye las medusas y los corales. Debido a que sólo el dorso inferior del organismo se ha fosilizado, no se conocen las suficientes características útiles del organismo para establecer una estructura adecuada.
 Bolsas:Fósiles como Pteridinium, conservados dentro de capas sedimentarias, se asemejan a «bolsas llenas de barro». La comunidad científica no ha llegado a un consenso sobre como interpretarlos.
Bolsas:Fósiles como Pteridinium, conservados dentro de capas sedimentarias, se asemejan a «bolsas llenas de barro». La comunidad científica no ha llegado a un consenso sobre como interpretarlos.
Organismos esponjosos:Tienen una apariencia «esponjosa» y son semejantes a un colchón inflable. En algunas ocasiones estas «esponjas» se rompieron o resquebrajaron antes de conservarse; estos ejemplares dañados ofrecen indicios valiosos para el proceso de reconstrucción.Estos organismos parecen conformar dos grupos, los organismos «rangeomorfos»,fractales, y los «erniettomorfos», más sencillos. Incluyendo en estos grupos los fósiles de los célebresCharniaySwartpuntia, este tipo es el más característico de los organismos ediacáricos, pero también el más difícil de situar dentro del árbol filogenético conocido. Al no disponer de boca, intestinos, órganos reproductivos ni conservarse ninguna prueba de su estructura interna, su existencia resulta muy extraña según los parámetros actuales. La hipótesis más aceptada es que podían succionar los nutrientes del agua a su alrededor porósmosis.
Ediacáricos «no ediacáricos»:Algunos organismos ediacáricos han conservado detalles más complejos, lo que ha permitido que sean interpretados como posibles formas primitivas de filos vivientes, excluyéndolos de algunas definiciones de los organismos ediacáricos. El fósil más antiguo de este tipo es el supuesto Vernanimalcula, bilateral, que según algunos especialistas es, en cambio, la parte interior de un saco de huevos o un acritarco.Una serie de fósiles conocidos como pequeños fósiles con caparazón están también representados en el periodo Ediacárico, siendo el más célebreCloudina, un pequeño fósil en forma de tubo que presenta marcas de depredación, sugiriendo que, aunque los predadores quizás no habrían sido muy comunes durante el Ediacárico, como mínimo existían.
 Icnofósiles: Los icnofósiles, también conocidos como pistas fósiles, son los restos fósiles de la actividad biológica que han quedado grabados en el sustrato, entre los que se encuentran las huellas, los cubiles, etc. Con la excepción de algunos escolitos muy simples, los únicos cubiles conocidos del periodo Ediacárico son horizontales y se localizan en la superficie o justo debajo. Estos cubiles implican que existían organismos móviles con cabezas, que probablemente habrían presentado una simetría bilateral.
Icnofósiles: Los icnofósiles, también conocidos como pistas fósiles, son los restos fósiles de la actividad biológica que han quedado grabados en el sustrato, entre los que se encuentran las huellas, los cubiles, etc. Con la excepción de algunos escolitos muy simples, los únicos cubiles conocidos del periodo Ediacárico son horizontales y se localizan en la superficie o justo debajo. Estos cubiles implican que existían organismos móviles con cabezas, que probablemente habrían presentado una simetría bilateral.

Probablemente su corta existencia sea debida bien a un cambio atmosférico y climático donde esta fauna no pudiese subsistir o bien porque la nueva fauna del cámbrico haya actuado como predadores sobre la fauna del ediacárico.
MORALEJA
Hay mucho misterio aun sobre esta fauna que aun no hemos descubierto por la falta de información,y dudo si se sabrá algún día pues la tierra ha evolucionado demasiado para aun encontrar restos que nos aclaren estas dudas.No obstante,yo pienso que fue claramente un intento fallido de la naturaleza para crear formas de vida que no resultó pero así se hacen mejores cosas,prueba y error.